关键信息
-
基因名
C6orf153
- 应用
-
别名
RRP36; C6orf153; HSPC253; Ribosomal RNA processing Protein 36 homolog
-
种属
Human
-
表达系统
E. coli
-
标签
GST-tag at N-terminal
-
纯度
Greater than 90% as determined by SDS-PAGE.
-
蛋白编号
Q96EU6
-
表达区间
1-259aa
-
氨基酸序列
MPGANYRAGAGAGAGARRPRGARDREEDGGGLEPAAVARDLLRGTSNMSFEELLELQSQVGTKTYKQLVAGNSPKKQASRPPIQNACVADKHRPLEMSAKIRVPFLRQVVPISKKVARDPRFDDLSGEYNPEVFDKTYQFLNDIRAKEKELVKKQLKKHLSGEEHEKLQQLLQRMEQQEMAQQERKQQQELHLALKQERRAQAQQGHRPYFLKKSEQRQLALAEKFKELKRSKKLENFLSRKRRRNAGKDRRHLPLSKE
-
分子量
56.2 kDa
-
内毒素
< 1.0 EU per μg protein as determined by the LAL method.
-
性状
Freeze-dried powder
-
缓冲液
PBS, pH7.4, containing 0.01% SKL, 1mM DTT, 5% Trehalose and Proclin300.
-
复溶方法
Reconstitute in ddH2O to a concentration of 0.1-0.5 mg/mL. Do not vortex.
- 个性化定制
-
稳定性测试
The thermal stability is described by the loss rate. The loss rate was determined by accelerated thermal degradation test, that is, incubate the protein at 37℃ for 48h, and no obvious degradation and precipitation were observed. The loss rate isless than 8% within the expiration date under appropriate storage condition.
-
保存条件 & 期限
Samples are stable for up to twelve months from date of receipt at -20℃ to -80℃. Store it under sterile conditions at -20℃ to -80℃. It is recommended that the protein be aliquoted for optimal storage. Avoid repeated freeze-thaw cycles.
-
运输条件
In general, recombinant proteins are supplied as lyophilized powder and shipped at ambient temperature. For bulk packages, the proteins are provided as frozen liquid and shipped with blue ice, unless otherwise requested by the customer.
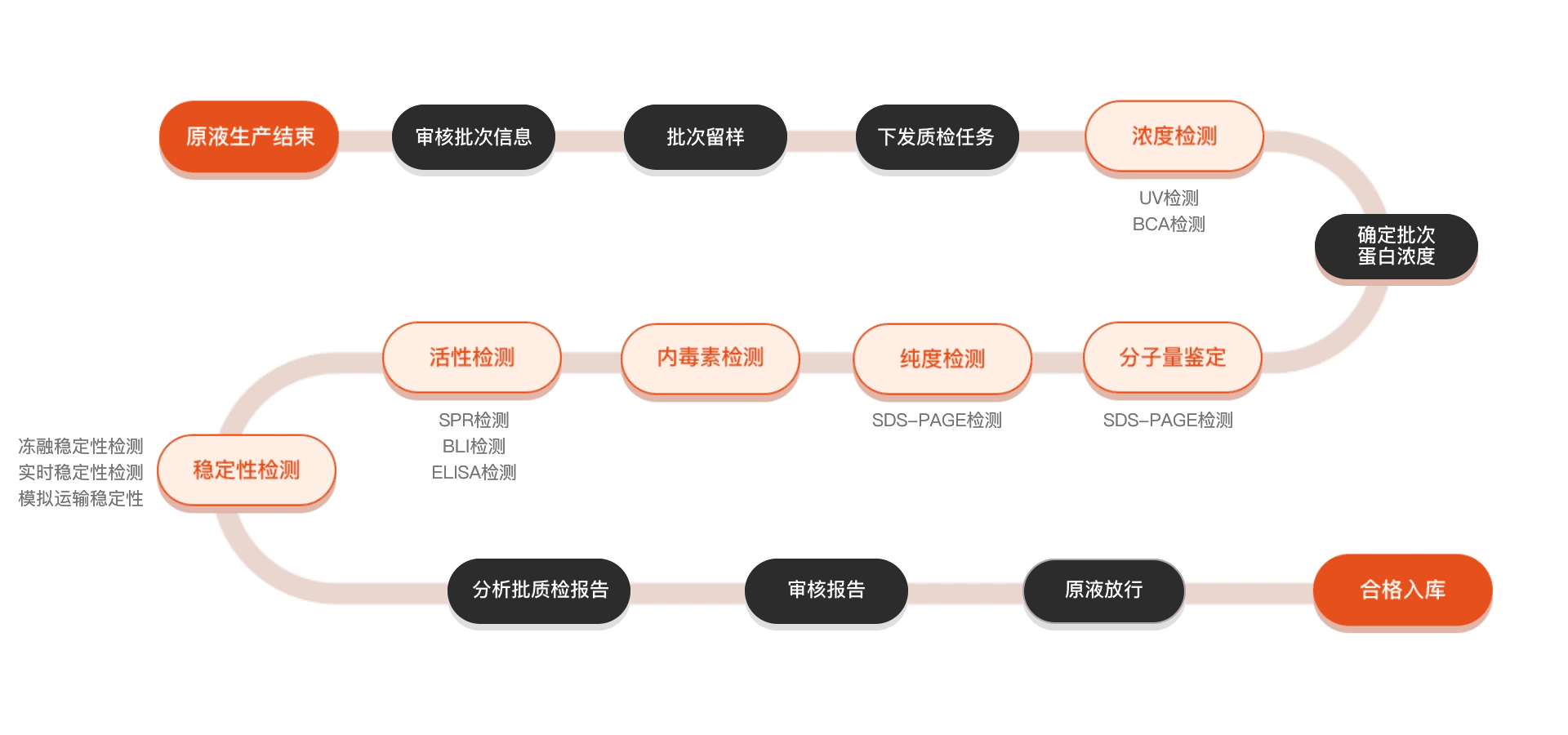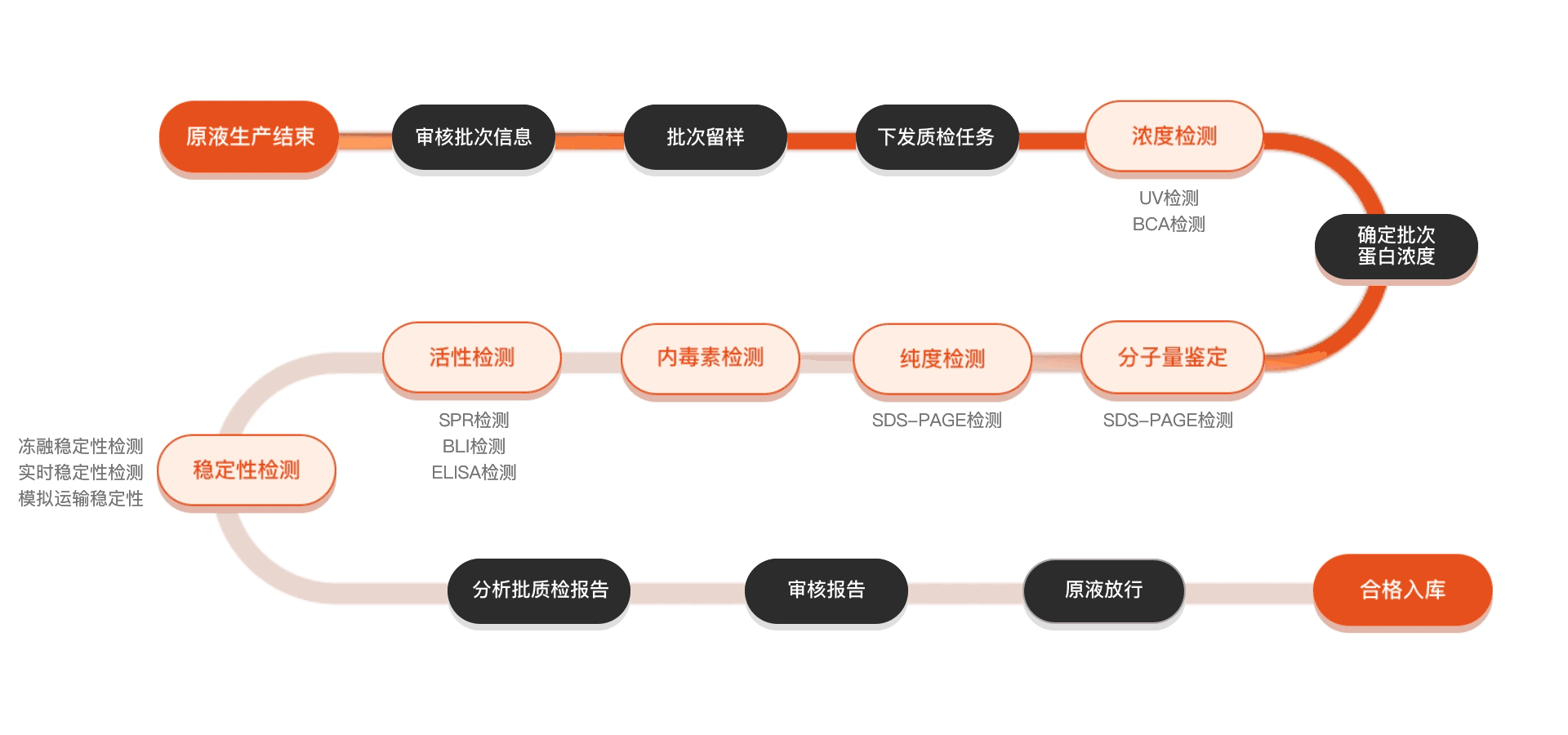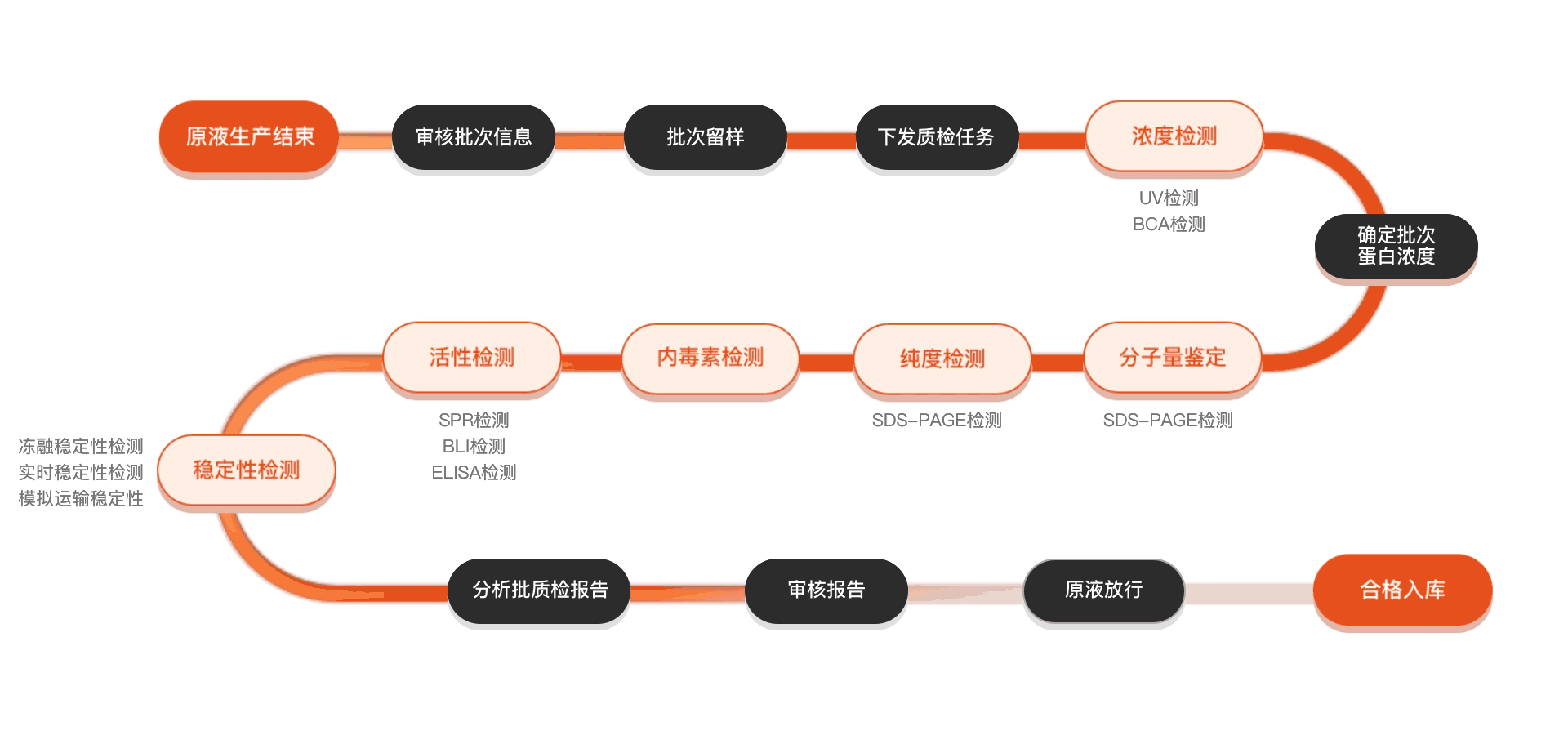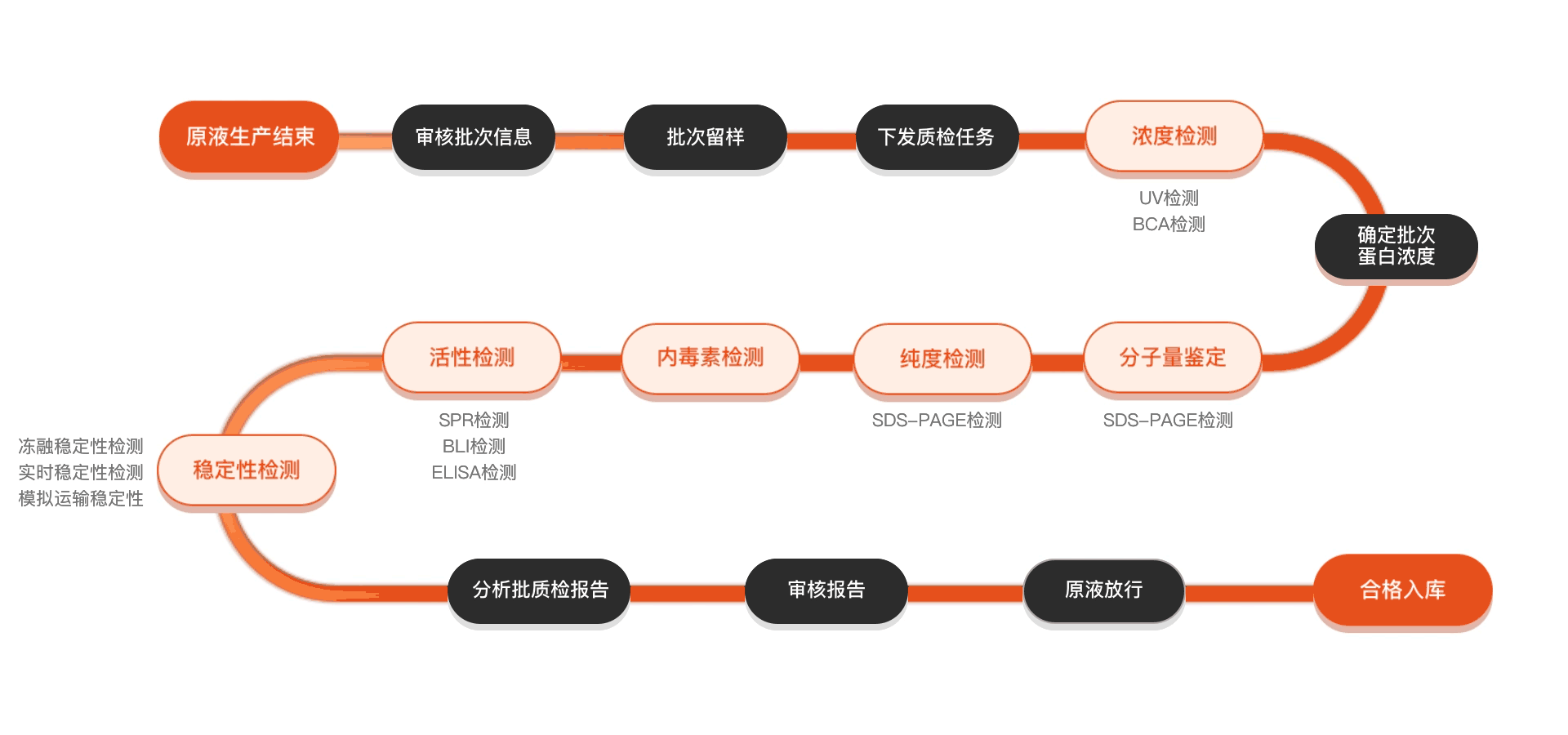
质检流程
相关产品






背景信息
C6orf153, also known as the uncharacterized protein product of the C6orf153 gene, has gained interest in recent years due to its potential roles in various biological processes. Located on chromosome 6, this gene is conserved across multiple species, suggesting a fundamental function. Initial studies indicated that C6orf153 may be involved in cellular mechanisms such as regulation of gene expression, cellular proliferation, and stress response. However, its precise biological roles and mechanisms of action remain largely unknown. Recent advancements in proteomics and molecular biology techniques have prompted researchers to explore the recombinant expression of C6orf153 to facilitate studies on its structure and function. By producing this protein in a heterologous system, scientists aim to conduct functional assays, structural studies, and protein interaction assays, which could elucidate its roles in cellular physiology and disease states. Understanding the functional implications of C6orf153 could uncover its relevance in health and disease, potentially impacting areas such as cancer research and therapeutic development. This research is crucial, as it addresses the gap in knowledge surrounding this enigmatic protein and could pave the way for novel biological insights and applications.